The simplest way to avoid infection is to prevent microorganisms gaining access …
The simplest way to avoid infection is to prevent microorganisms gaining access to the body. The skin has an external coating of dead cells (cuticle) that, when intact, is impermeable to most infectious agents as very few pathogens are capable of penetrating the thick stratified squamous epithelium of the skin (and lower urinary tract).
Pathogens can invade the body if a breach occurs in the barriers …
Pathogens can invade the body if a breach occurs in the barriers formed by the skin and mucus membranes, for example a wound, they must be detected and destroyed by cellular and humoral means. The cells involved in the cellular response to a wound are mast cells, macrophages, granulocytes, and monocytes.
The innate response to bacterial infection lies in its first-response role of …
The innate response to bacterial infection lies in its first-response role of detection of a foreign organism. By using the tools of Pattern-Recognition Receptors (PRRs), the innate response flags up problems while the adaptive response gets itself organized.
Because viruses invade host cells to take over a host's cellular machinery, …
Because viruses invade host cells to take over a host's cellular machinery, the innate system has a more difficult time detecting viruses as foreign agents. However, there is a give-away element of the viral attack that the innate system can recognize: the double-stranded RNA (dsRNA) produced by a virus in its replication phase. Because mammalian cells only ever produce single-stranded RNA, the presence of dsRNA signals a foreign intruder. dsRNA can be detected by TLR-3R on the cell surface or intracellularly by the presence of dsRNA-dependent protein kinase.
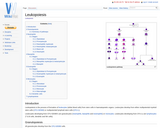
Leukopoiesis is the process of formation of leukocytes (white blood cells) from …
Leukopoiesis is the process of formation of leukocytes (white blood cells) from stem cells in haematopoietic organs. Leukocytes develop from either multipotential myeloid stem cells (CFU-GEMM) or multipotential lymphoid stem cells (CFU-L).
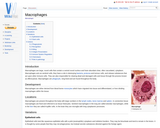
Macrophages are large, round cells that contain a central round nucleus and …
Macrophages are large, round cells that contain a central round nucleus and have abundant clear, often vacuolated, cytoplasm. Macrophages acts as sentinel cells; they have a role in destroying bacteria, protozoa and tumour cells, and release substances that act upon other immune cells. They are also responsible for clearing dead and damaged cells and tissue through the process known as efferocytosis. Macrophages are phagocytic, long lived and are found throughout the body.
T-cells rely on Major Histocompatability Complexes (MHC), which are molecules manufactured within …
T-cells rely on Major Histocompatability Complexes (MHC), which are molecules manufactured within cells for the purpose of presenting antigen fragments so that they can be detected by the immune system. MHC has evolved to form two classes for antigen presentation: MHC I presents digested fragments from antigen in cellular cytoplasm, and MHC II presents digested fragments from antigen in the tissue fluid (extracellular). MHC I tends to bind slightly smaller peptides (~9 amino acids) than MHC II (~15 amino acids).
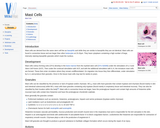
Mast cells are derived from the same stem cell line as basophils …
Mast cells are derived from the same stem cell line as basophils and while they are similar to basophils they are not identical. Mast cells are found in connective tissue and are larger than other leukocytes at 15-20ľm. They have cytoplasm containing a high number of large, intensely staining basophilic granules which mask the nucleus.
Monocytes are the largest leukocytes. They have a large indented nucleus, few …
Monocytes are the largest leukocytes. They have a large indented nucleus, few granules and constitute about 5% of circulating leukocytes. Monocytes are precursors to a number of cells that make up the mononuclear phagocytic system. Developing in the bone marrow, monocytes migrate into the circulation where, after approximately three days, they migrate into tissues and differentiate.
NK cells can be classified as lymphocytes because they are capable of …
NK cells can be classified as lymphocytes because they are capable of recognising antigen, however they are more often associated with the innate immune response. They target cells by monitoring MHC production, which is expressed by healthy cells to present antigen to T-cells. Low MHC levels can be used as a marker for a cell whose machinery is compromised by a replicating virus. When MHC levels drop, it acts as a danger signal to the NK cells, which then release enzymes to kill the infected cells.
Neutrophils represent up to 70% of all leukocytes in the blood stream …
Neutrophils represent up to 70% of all leukocytes in the blood stream (in humans there are approximately 4.4 million neutrophils / millilitre of blood) and are distinguished by their irregular multi-lobed nucleus and indistinct granular appearance. They are 10-12ľm and circulate in the blood for minutes to hours (average 6-8 hours) and lasting for 1-2 days in tissue. Although the most abundant leukocyte in the blood, the vast majority of neutrophils are found in the bone marrow (5x more) mostly as functionally immature precursor cells, although this varies between species with mice having a large pool of functional neutrophils in their bone marrow.
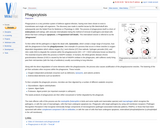
Phagocytosis is a very primitive system of defence against infection, having even …
Phagocytosis is a very primitive system of defence against infection, having even been shown to exist in invertebrates and single cell organisms. The discovery was made in starfish larvae by Elle Metchnikoff who subsequently won the Nobel Prize for Medicine or Physiology in 1908. The process of phagocytosis itself is a form of endocytosis (cell eating), with vesicular internalisation being the method of removal of pathogens and dead cells (those that have undergone apoptosis, or Programmed Cell Death). This internalised vesicle is referred to as the "phagosome".
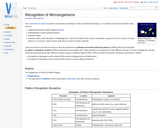
The innate immune system recognises components of pathogens which are intrinsically foreign …
The innate immune system recognises components of pathogens which are intrinsically foreign (i.e. not present on normal mammalian cells), such as Lipolysaccharides, Peptidoglycans and D-isoform amino acids.
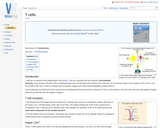
T cells are so named as they differentiate in the thymus. They …
T cells are so named as they differentiate in the thymus. They are long lived and are involved in cell mediated immunity. They represent 60-80% of the circulating lymphocytes and all express the markers CD2, CD3 and CD7 as well as having T cell receptors (TCR). Each T cell has 30,000 TCRs each of which is identical and recognises antigens and major histocompatability complex (MHC) II.
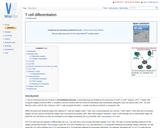
T cells are long lived and are involved in cell mediated immunity. …
T cells are long lived and are involved in cell mediated immunity. Functionally they are divided by the expression of CD4+ or CD8+ markers. CD4+ T helper cells recognise antigens bound to MHC II complexes and are involved with the control of intracellular and extracellular pathogens; they can interact with CD8+, NK and dendritic cells or with B cells. Cytotoxic CD8+ T cells recognise the MHC I complex and destroy infected or neoplastic cells.
Thrombocytes are small anuclear fragments of megakaryocytes. They are membrane bound portions …
Thrombocytes are small anuclear fragments of megakaryocytes. They are membrane bound portions of the megakaryocyte cytoplasm and have a finely granular cytoplasm; they are much smaller than other blood cells at 2-3ľm and have a lifespan of around 10 days in the circulation.
As with erythrocytes, thrombocytes are derived from multipotential myeloid stem cells (CFU-GEMM). …
As with erythrocytes, thrombocytes are derived from multipotential myeloid stem cells (CFU-GEMM). In the bone marrow CFU-GEMM cells differentiate into the megakaryocyte precursor cell the megakaryocyte CFU (CFU-Meg), under the influence of cytokines CFU-CSF and IL-3. Unlike other blood cells which undergo mitosis in the first few developmental stages, once the CFU-GEMM has differentiated into the CFU-Meg there is no further mitosis. The CFU-Meg then develops into the megakaryoblast.
No restrictions on your remixing, redistributing, or making derivative works. Give credit to the author, as required.
Your remixing, redistributing, or making derivatives works comes with some restrictions, including how it is shared.
Your redistributing comes with some restrictions. Do not remix or make derivative works.
Most restrictive license type. Prohibits most uses, sharing, and any changes.
Copyrighted materials, available under Fair Use and the TEACH Act for US-based educators, or other custom arrangements. Go to the resource provider to see their individual restrictions.